Research Article
Effect of common food additives on mast cell activation

Carena MP, Mariani ML, Ordóñez A and Penissi AB*
Instituto de Histología y Embriología “Dr. Mario H. Burgos” (IHEM-CONICET), Facultad de Ciencias Médicas, Universidad Nacional de Cuyo, Argentina
*Address for Correspondence: Alicia Beatriz Penissi, Professor, PhD, Instituto de Histología y Embriología “Dr. Mario H. Burgos” (IHEM-CONICET), Facultad de Ciencias Médicas, Universidad Nacional de Cuyo. Casilla de Correo 56. (5500) Mendoza, Argentina, Tel: 00-54-0261-4135000; 2670/2672/2673; Fax: 00-54-261-4494117; Email: [email protected]
Dates: Submitted: 03 January 2019; Approved: 16 January 2019; Published: 17 January 2019
How to cite this article: Carena MP, Mariani ML, Ordóñez A, Penissi AB. Effect of common food additives on mast cell activation. Insights Clin Cell Immunol. 2019; 3: 001-005. DOI: 10.29328/journal.icci.1001007
Copyright License: © 2019 Carena MP, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.

Abstract
Mast cells play a central role in the genesis and modulation of allergic and inflammatory responses. The general aim of the present work was to study the interaction between mast cells and the most common additives approved for use in foods. Dose-response studies about the effect of the main food additives (tartrazine, sodium bisulphite and sodium benzoate) on mast cell degranulation were carried out. Rat peritoneal mast cells were incubated with: 1) buffer solution or 2) stimulus. The stimuli were tartrazine, sodium benzoate, sodium bisulphite and the calcium ionophore A23187. A23187 was used as a reference mast cell secretagogue. Different doses and combinations of food additives were used. The viability of the mast cells was evaluated with trypan blue. In the incubation solutions, the release of β-hexosaminidase was quantified by colorimetric reaction and ELISA plate reader. The remaining β-hexosaminidase concentration (not released) was studied in the cells after the incubations, and morphology of the mast cells was analyzed by light microscopy with toluidine blue stain. The food additives tartrazine, sodium benzoate and sodium bisulphite did not stimulate the release of β-hexosaminidase from mast cells at any of the concentrations used. In contrast, tartrazine at concentrations of 0.1 μM and 1 μM, and sodium benzoate and sodium bisulphite at concentrations of 0.1 μM, 1 μM, 10 μM and 100 μM, significantly inhibited the basal release of β-hexosaminidase from mast cells. Considering these findings, we decided to determine the effect of these additives on the degranulation of mast cells induced by the calcium ionophore A23187. Sodium bisulphite inhibited mast cell activation induced by the calcium ionophore A23187 in this experimental model. The present study demonstrates that food additives of usual permitted use do not stimulate basal degranulation of mast cells in an in vitro model of peritoneal mast cells and that the additive sodium bisulphite inhibit mast cell activation induced by intracellular calcium increase. This food additive could represent an interesting alternative in the prevention of pathologies mediated by mast cells, as well as in the field of nutritional biochemistry.
Introduction
Mast cells are specialized connective tissue cells that play a central role in the genesis and modulation of allergic and inflammatory responses [1-4].
Considering the allergenic properties described by some authors of the main food additives allowed (tartrazine, sodium bisulphite and sodium benzoate) [5-7], our working hypothesis suggests that these food additives stimulate degranulation of mast cells.
The general aim of the present work was to study the interaction between mast cells and the most common additives approved for use in foods.
Material and Methods
Assayed compounds
The food additives tartrazine, sodium benzoate and sodium bisulphite, as well as the calcium ionophore A23187 were purchased from Sigma.
Animals
Male Wistar adult rats weighing approximately 300 to 500 g, infection free and maintained under a 12-hour dark/light cycle in a temperature-controlled room (24-25ºC) with free access to drinking water and laboratory food, were used for the study. All animal experiments were carried out according to the standards included in the Guide for the Care and Use of Laboratory Animals (published by the National Academy of Science, National Academy Press, Washington, D.C.), and approved by the Institutional Committee for Care and Use of Laboratory Animals (CICUAL, Facultad de Ciencias Médicas, Universidad Nacional de Cuyo, Mendoza, Argentina).
Isolation and purification of rat mast cells
Mast cells were obtained by peritoneal lavage with physiological solution and purified in a discontinuous gradient of sterile Percoll solution. The purity of the cells was evaluated by staining with 1% toluidine blue. For the tests, preparations with purity of 95-100% were used. The viability of the mast cells was evaluated with trypan blue.
General protocol
The purified mast cells were incubated with: 1) buffer solution or 2) stimulus. The stimuli were tartrazine, sodium benzoate, sodium bisulphite and the calcium ionophore A23187. A23187 was used as a reference mast cell secretagogue. Different doses and combinations of food additives were used (these doses include those commonly used in food and those referred by the literature). The viability of the mast cells was evaluated with trypan blue. In the incubation solutions, the release of β-hexosaminidase was quantified by colorimetric reaction and ELISA plate reader (the release of this enzyme is used as a mast cell degranulation marker). The remaining β-hexosaminidase concentration (not released) was studied in the cells after the incubations. Morphology of the mast cells was analyzed by light microscopy after experiments.
Quantification of β-hexosaminidase
β-hexosaminidase levels, both in the incubation solutions and in the remaining cells, was measured by colorimetric reaction and ELISA plate reader.
Light microscopy with blue toluidine stain
Mast cells were fixed in 2% glutaraldehyde. After 2 h in the fixative, cell suspensions were stained with toluidine blue (0.1% w/v, pH 3.0), placed between slides and coverslides, and examined under an Olympus CH-2 microscope. Percentage of mast cells exhibiting degranulation was quantitated at a magnification of X 400. Mast cell activation was defined as the presence of extruded granules close to the surface of the cell in question or staining of about half or less of the cell with toluidine blue.
Statistical treatment
The results were analyzed using analysis of variance type 1 (ANOVA-1) followed by the Tukey-Kramer test. P <0.05 was considered statistically significant.
Results
The food additives tartrazine, sodium benzoate and sodium bisulphite did not stimulate the release of β-hexosaminidase from mast cells at any of the concentrations used (Figures 1-4). In contrast, tartrazine at concentrations of 0.1 μM and 1 μM, and sodium benzoate and sodium bisulphite at concentrations of 0.1 μM, 1 μM, 10 μM and 100 μM, significantly inhibited the basal release of β-hexosaminidase from mast cells (Figures 1-4). Higher concentrations of the additives did not inhibit this release, probably due to cell death.
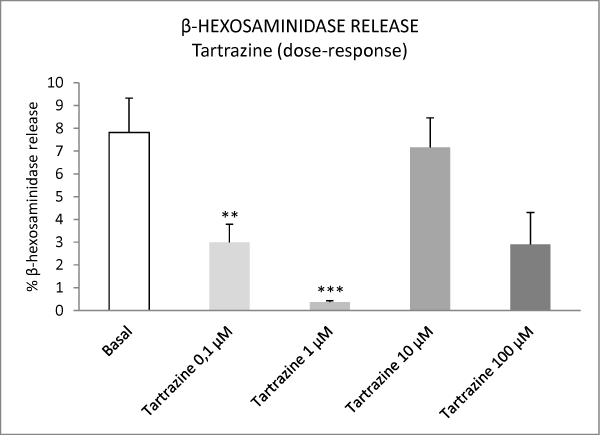
Figure 1: Effect of varying concentrations of tartrazine on basal β-hexosaminidase release from peritoneal mast cells. Results are expressed as percentage release of β-hexosaminidase. Values are presented as means ± S.E.M. **P<0.01 versus basal; ***P<0.001 versus basal.
Figure 2: Effect of varying concentrations of sodium benzoate on basal β-hexosaminidase release from peritoneal mast cells. Results are expressed as percentage release of β-hexosaminidase. Values are presented as means ± S.E.M. **P<0.01 versus basal; ***P<0.001 versus basal.
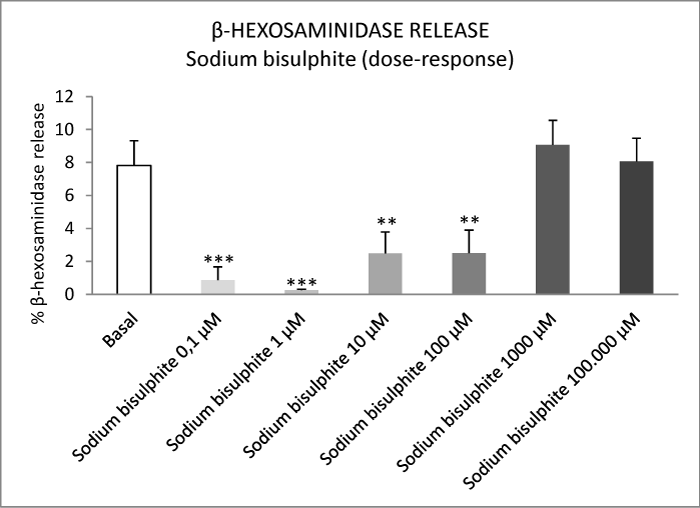
Figure 3: Effect of varying concentrations of sodium bisulphite on basal β-hexosaminidase release from peritoneal mast cells. Results are expressed as percentage release of β-hexosaminidase. Values are presented as means ± S.E.M. **P<0.01 versus basal; ***P<0.001 versus basal.
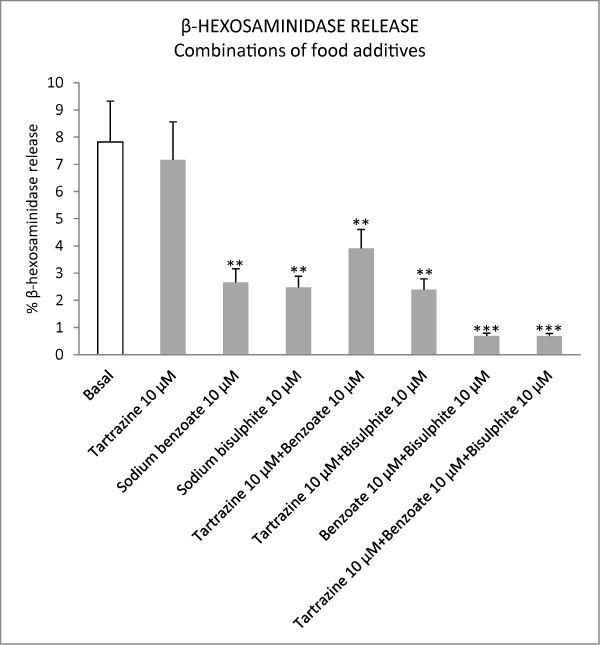
Figure 4: Effect of different combinations of food additives on basal β-hexosaminidase release from peritoneal mast cells. Results are expressed as percentage release of β-hexosaminidase. Values are presented as means ± S.E.M. **P<0.01 versus basal; ***P<0.001 versus basal.
The inhibitory action of the food additives was not accompanied by changes in cell viability. The trypan blue-exclusion test indicated a viability of greater than 80% at inhibitory concentrations, and lower than 80% at not inhibitory doses (data not shown).
Considering that some of the commonly used food additives (tartrazine, sodium benzoate and sodium bisulphite in concentrations of 0.1 μM, 1 μM) did not stimulate but inhibited the degranulation of peritoneal mast cells, we decided to determine the effect of these additives on the degranulation of mast cells induced by the calcium ionophore A23187. Sodium bisulphite (alone and combined with sodium benzoate and/or sodium bisulphite) inhibited mast cell activation induced by the calcium ionophore A23187 in this experimental model. The results obtained are shown in figure 5.
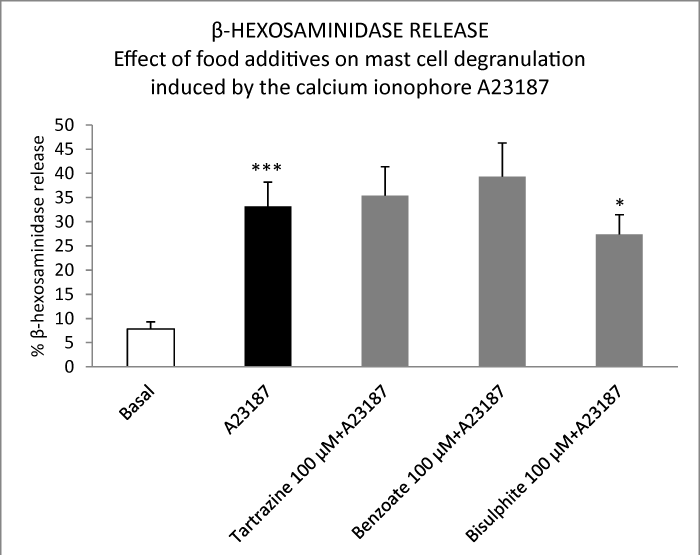
Figure 5: Effect of food additives on mast cell β-hexosaminidase release induced by the calcium ionophore A23187. Results are expressed as percentage release of β-hexosaminidase. Values are presented as means ± S.E.M. ***P<0.001 versus basal; *P<0.05 versus A23187.
Our biochemical results about β-hexosaminidase release are consistent with those obtained by light and microscopy (Figure 6).
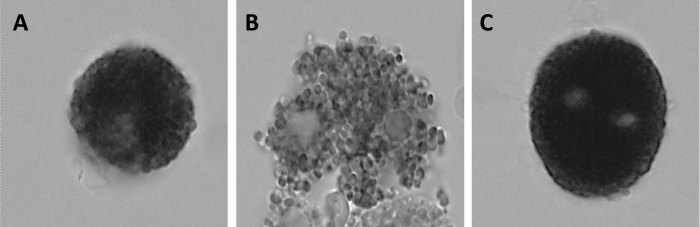
Figure 6: Light-microscopic photographs of peritoneal mast cells (toluidine blue stain). A: basal. The cytoplasm is dominated by closely packed secretory granules. B: A23187. Degranulating mast cell may be seen. C: sodium bisulphite+A23187. The morphology of bisulphite+A23187-treated cells shows a lower degree of degranulation than that of secretagogue samples. X 600.
Discussion and Conclusions
The present study shows that the commonly used food additives (tartrazine, sodium benzoate and sodium bisulphite) do not stimulate basal degranulation of mast cells in an in vitro model of peritoneal mast cells. By the contrary, these additives inhibit the basal release of β-hexosaminidase from mast cells at low concentrations. This means that some physiological effects of mast cells could be affected by these food additives.
However, sodium bisulphite inhibits mast cell activation induced by the calcium ionophore A23187 in this same experimental model. It is the first time that a stabilizing action at mast cell level has been demonstrated for sodium bisulphite.
The mechanism of the inhibitory action of sodium bisulphite would be related to the ability of this compound to react with carbonyl compounds and to inhibit FcεRI receptors [8,9]. However, further research is needed to explain the exact molecular mechanisms of these actions.
This compound could represent interesting alternative in the treatment and / or prevention of pathologies mediated by mast cells, as well as in the field of nutritional biochemistry.
References
- Galli SJ, Grimbaldeston M, Tsai M. Immunomodulatory mast cells: negative, as well as positive, regulators of immunity. Nature. 2008; 8: 478-486. Ref.: https://goo.gl/iKFxPo
- Kawakami T, Ando T, Kimura M, Wilson BS, Kawakami Y. Mast cells in atopic dermatitis. Curr Opin Immunol. 2009; 21: 666-678. Ref.: https://goo.gl/YwzDmq
- Hofmann AM, Abraham SN. New roles for mast cells in modulating allergic reactions and immunity against pathogens. Curr Opin Immunol. 2009; 21: 679-686. Ref.: https://goo.gl/PbjQjp
- Stone KD, Prussin C, Metcalfe DD. IgE, mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 2010; 125: S73-S80. Ref.: https://goo.gl/CoLbdp
- Freedman BJ. Asthma induced by sulphur dioxide, benzoate and tartrazine contained in orange drinks. Clin Allergy. 1977; 7: 407-415. Ref.: https://goo.gl/kbn8zq
- Safford RJ, Goodwin BF. The effect of tartrazine on histamine release from rat peritoneal mast cells. Int J Immunopharmacol. 1984; 6: 233-240. Ref.: https://goo.gl/3rKhE1
- Collado CR, Hochman DJ, Goldblum RM, Brooks EG. Effect of sodium sulfite on mast cell degranulation and oxidant stress. Ann Allergy Asthma Immunol. 2006; 96: 550-556. Ref.: https://goo.gl/UT2xeR
- Lai MD, Xu YH. Effects of bisulphite and sulfite on Fc and Con A receptors of rat pulmonary macrophages. Chin Med J.1988; 101:357-360. Ref.: https://goo.gl/n15rbN
- Maeda-Yamamoto M1, Ema K, Monobe M, Tokuda Y, Tachibana H. Epicatechin-3-O-(3″-O-methyl)-gallate Content in Various Tea Cultivars (Camellia sinensis L.) and Its in Vitro Inhibitory Effect on Histamine Release. J Agric Food Chem. 2012; 60: 2165-2170. Ref.: https://goo.gl/nbDrLh